

2024年7月17日,曾出演《卧虎藏龙》、《唐伯虎点秋香》的著名演员郑佩佩去世。据报道早在2019年,郑佩佩被诊断为神经退行性非典型帕金森综合征——非正式名称皮质基底节变性(Corticobasal degeneration, CBD)。CBD属于tau病理,帕金森属于α-syn病理,CBD诊断比较困难,目前也无特效药。郑佩佩选择将大脑捐赠给Brain Support Network(BSN),遗体捐给香港大学以供医学研究。
一代女侠,她的伟大不仅体现在艺术创作上,是无私的奉献精神的体现,是对人类医学研究巨大的贡献,更是侠义精神的延续。侠肝义胆与不凡的气节让人深深触动。普瑞麦迪公司缅怀郑佩佩女士的离世,特此汇总了FIDA技术在近期发表的一系列关于神经退行性疾病研究的案例,我们相信卓越的科研人员,使用前沿的研究工具,一定能早日攻克神经退行性疾病这个恶魔。
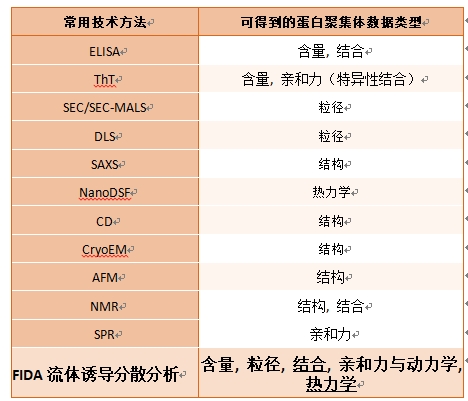
FIDA多维表征蛋白聚集体:含量, 粒径, 结合, 亲和力与动力学, 热力学数据
神经退行性疾病是神经元结构或功能逐渐丧失甚至死亡,继而导致功能性障碍的一类疾病。帕金森氏症、阿尔茨海默氏症、亨廷顿氏症,就是典型的神经退行性疾病。
神经退行性疾病病理标志是错误折叠蛋白纤维聚集体的形成。然而表征蛋白聚集体的多种参数与寻找能够结合蛋白聚集体的新型分子面临着诸多挑战。传统的相关检测技术受限于检测灵敏度、准确性或通量等问题,在神经退行性疾病的研究中存在很多局限性,往往需要多种方法结合才能得出理想数据,但有限的样本数量很难支持众多的实验类型。
FIDA技术由于在底层物理原理上的优势,可以解决现有方法无法克服的一系列问题,例如结合质量变化不敏感,蛋白聚集体沉淀,结合构象变化小,Labelfree信号弱难以检测,样品量小需要在细胞裂解液中检测等问题。FIDA可以在一次实验中得到目前常用方法得到的多个表征参数。
下面 ,将通过案例详细解析FIDA流体诱导分散技术如何表征蛋白聚集体尺寸, 结合特异性, 亲和力, 热力学等数据。
FIDA在淀粉样蛋白疾病诊断工具肽开发的应用
蛋白质聚集与淀粉样蛋白疾病的发展有关,如阿尔茨海默病和亨廷顿病。尽早发现聚集物非常重要,这是早期诊断的先决条件。本文提出了一种从早期聚集监测蛋白原纤维大小的方法。研究者设计了一种名为FibrilPaint1的肽联合FIDA技术,它能特异性地识别来自疾病的各种原纤维,但不能识别它们的单体,这个特性使FibrilPaint1成为淀粉样蛋白疾病诊断应用的新工具。
筛选FibrilPaint肽,FIDA快速检测能与Tau和HttEx1Q44单体蛋白与原纤维结合的多肽
最初,研究者筛选了四种FibrilPaint肽,以直接检测能否结合Tau和HttEx1Q44原纤维。使用FIDA技术,对FibrilPaint肽进行荧光标记,从荧光信号中测量多肽的绝对水动力半径(Rh)变化。
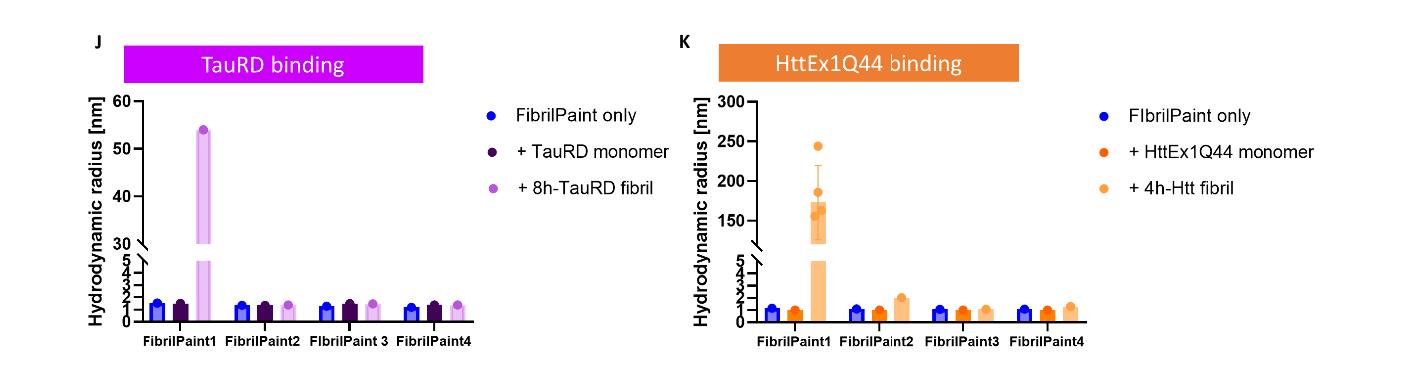 图1,FibrilPaint1-4都抑制了单体聚集,但FibrilPaint1与Tau和HttEx1Q44原纤维结合,而且不与其单体结合
图1,FibrilPaint1-4都抑制了单体聚集,但FibrilPaint1与Tau和HttEx1Q44原纤维结合,而且不与其单体结合
四种FibrilPaint肽都能抑制TauRD的聚集体的产生,但FibrilPaint1不仅有抑制能力,由于它与TauRD亲和力很高,可作为蛋白聚集体的非共价标签。这使得FibrilPaint1同时具备的抑制聚集体产生,与作为TauRD的聚集体的标签工具的能力,所以科学家继续利用FIDA与FibrilPaint1开发可以诊断TauRD相关疾病的工具与方法。
利用FIDA确认在细胞裂解液和聚集体中FibrilPaint1对纤维结合的特异性
文章中研究了 FibrilPaint1 是否在不受其他生物分子或细胞成分干扰的情况下结合纤维。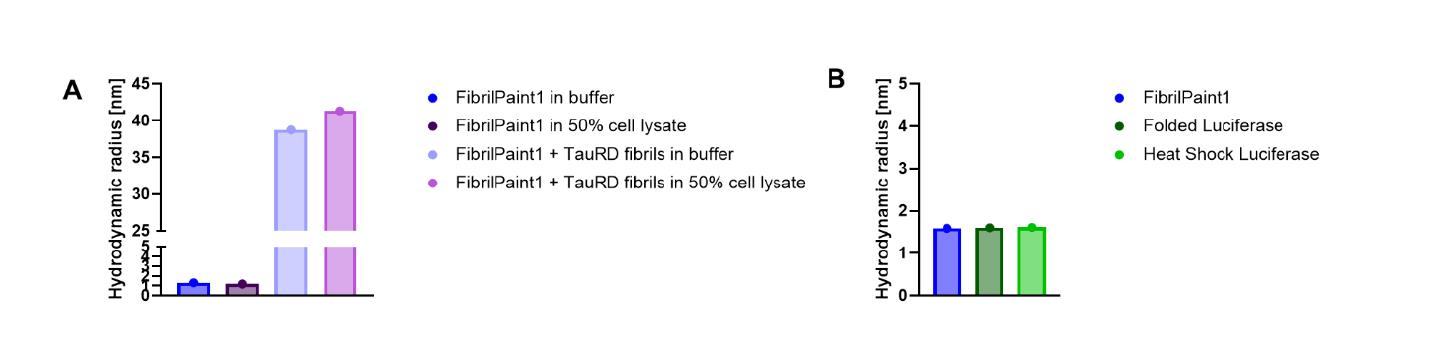 图2,FibrilPaint1对淀粉样蛋白聚集体具有特异性
图2,FibrilPaint1对淀粉样蛋白聚集体具有特异性
通过加入大肠杆菌细胞裂解液,测试了其在大量其他细胞蛋白存在下特异性识别 TauRD 纤维的能力。在 50% 细胞裂解液中,以 1:100 的比例将预先形成的 20 小时 TauRD 纤维与 FibrilPaint1 一起孵育。在这些条件下,FibrilPaint1 的尺寸增加到 39 nm,这与在缓冲液中测量到的相同纤维的 41 nm Rh 值一致(图3A)。当单独在细胞裂解液中孵育 FibrilPaint1 时,其尺寸与单独的 FibrilPaint1 保持不变(图2A)。因此,FibrilPaint1 能够在复杂的细胞混合物中特异性识别纤维。
接下来,测试了 FibrilPaint1 与聚集体结合是否需要纤维状结构。使用萤火虫荧光素酶作为非淀粉样蛋白聚集体的标准参照。萤火虫荧光素酶是一种球状的 61 KDa 蛋白,其 Rh 为 3.4 nm,在热冲击变性时形成无定形聚集体。以 1:100 的比例将 FibrilPaint1 与热冲击萤火虫荧光素酶一起孵育。在 FIDA 测量中,FibrilPaint1 的 Rh 不受萤火虫荧光素酶聚集体存在的影响(图2B)。这表明 FibrilPaint1 不与无定形的萤火虫荧光素酶聚集体结合。综上所述,这些数据表明 FibrilPaint1 在不受其他生物分子干扰的情况下,特异性结合淀粉样蛋白纤维。
FIDA快速表征Tau蛋白和Htt蛋白纤维绝对粒径的动力学变化
接下来,研究者将FIDA获得的与单颗粒EM获得的纤维粒径大小进行比较。使用阴性染色EM成像来表征聚集24小时后的TauRD原纤维和聚集6小时后的HttEx1Q44原纤维的形状大小。采用ThT和FIDA平行测定两种蛋白聚集的变化,ThT定性提供淀粉样蛋白原纤维存在的信号。FIDA量化提供了纤维长度在早期聚集期随时间的发展绝度粒径 Rh大小,与负染色EM成像数据一致。
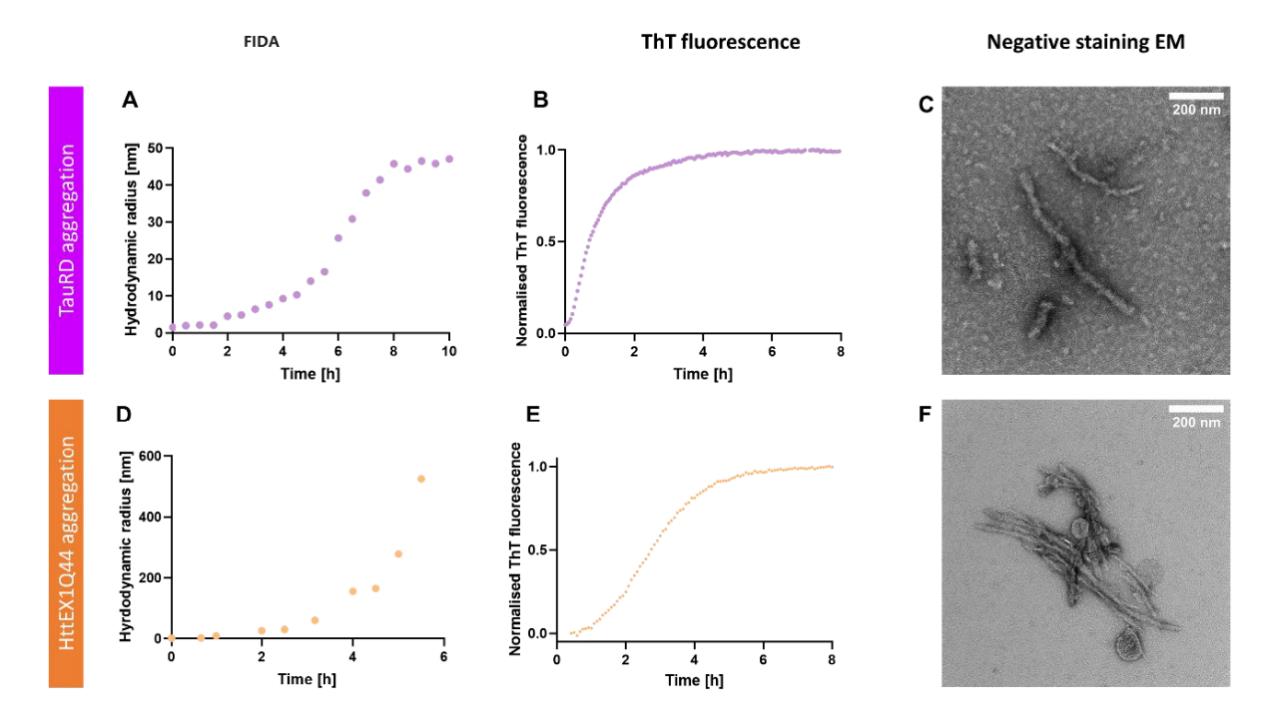 图3,TauRD (A)和HttEx1Q44 (D)聚集物的流体动力半径用FIDA测定。在平行实验中,TauRD和HttEx1Q44聚集物用ThT测定(B, E)监测。TauRD的最终产物在24小时后(C)和6小时后(F)的电镜染色为阴性,Tau-RD聚集,HttEx1Q44聚合成簇状
图3,TauRD (A)和HttEx1Q44 (D)聚集物的流体动力半径用FIDA测定。在平行实验中,TauRD和HttEx1Q44聚集物用ThT测定(B, E)监测。TauRD的最终产物在24小时后(C)和6小时后(F)的电镜染色为阴性,Tau-RD聚集,HttEx1Q44聚合成簇状
FIDA确定FibrilPaint1结合特异性——监测患者来源的原纤维
FibrilPaint1特异性结合淀粉样蛋白原纤维,它可能具有诊断患者蛋白原纤维的潜力。这就要求FibrilPaint1能够识别患者来源的原纤维。
接下来,研究者评估FibrilPaint1识别多种Tau病变患者源性原纤维的能力,采用了三种不同Tau病变的离体Tau原纤维,分别从诊断为CBD、FTD和AD的已故患者中纯化得到。
用阴性染色EM对纯化的原纤维进行成像,以确认典型的疾病特异性原纤维形状。随后,进行FIDA分析以确定患者来源的Tau原纤维的大小(A)。重组TauRD形成的原纤维平均Rh为54 nm。从诊断为AD的患者中测量的原纤维的表观平均Rh为40 nm,测量值之间的变化很小(A)。对于CBD,观察到的Rh为140 nm(图A)。来自FTD的原纤维是一组均匀的蛋白聚集体,平均 Rh为60 nm(A)。这些数据表明,FibrilPaint1适合与源自原材料的蛋白原纤维相互作用并表征。
最后用这些原纤维中的重复元素结合EM数据,来确定它们的平均长度。PHF原纤维的平均长度为2.7倍重复,相当于430 nm。这与fibrilpaint1染色原纤维在FIDA实验中观察到的420 nm一致。因此,联合FibrilPaint1和FIDA是确定病理原纤维长度的合适方法
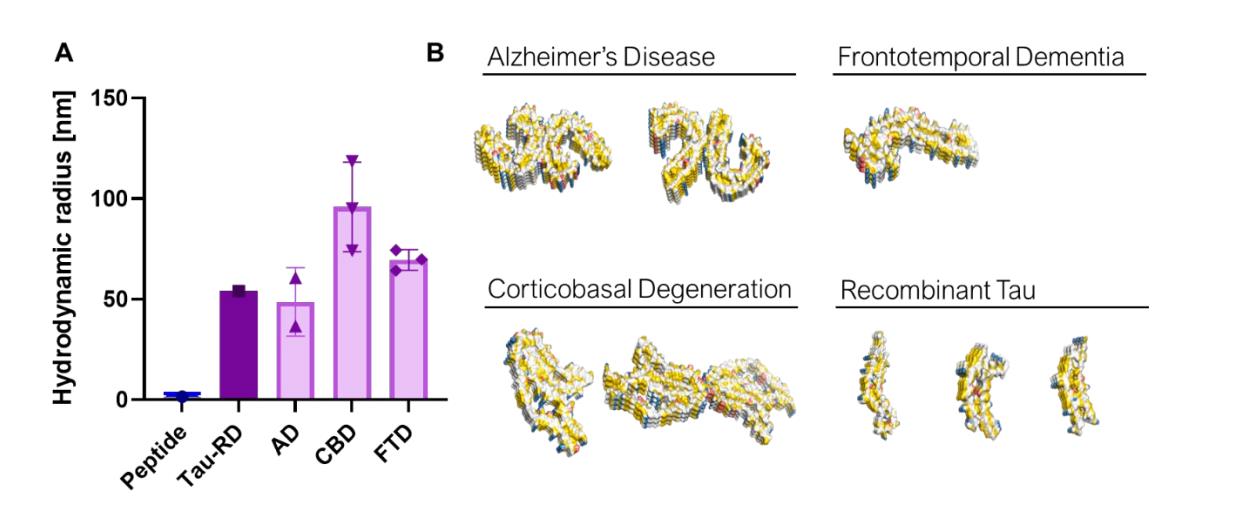
图4,FIDA检测FibrilPaint1与不同来源Tau蛋白聚集体的结合特异性
由于FibrilPaint1可以准确识别多种不同结构Tau病变患者源性原纤维,所以基于FIDA开发了一种采用FibrilPaint1标记tau原纤维,并且测量原纤维的大小的淀粉样蛋白疾病诊断工具。
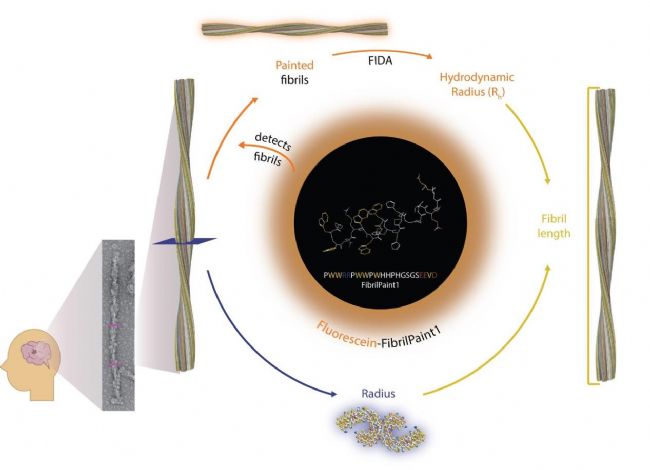
图5,基于FIDA的FibrilPaint1探测淀粉样蛋白原纤维的方法
通过FibrilPaint1与不同来源的Tau蛋白的强结合能力,可以检测淀粉样蛋白原纤维的存在。通过FibrilPaint1加上荧光素标签,使结合了FibrilPaint1的原纤维被染色(橙色途径),从而诊断淀粉样疾病的发生。然后,染色后的纤维可以用FIDA分析得到蛋白聚集体的水动力半径(Rh),其中如聚集体的结构是已知结构中的,可以进一步计算出确定纤维的半径(蓝色途径),从而更准确的诊断Tau蛋白相关疾病与发生机制。
来源:Fibril Paint to detect Amyloids and determine Fibril Length Júlia Aragonès Pedrola, Françoise A. Dekker, Tommaso Garfagnini, Guy Mayer, Margreet B. Koopman, Menno Bergmeijer, Friedrich Förster, Jeroen J. M. Hoozemans, Henrik Jensen, Assaf Friedler, Stefan G. D. Rüdiger bioRxiv 2023.10.13.562220; doi: https://doi.org/10.1101/2023.10.13.562220
FIDA表征淀粉样蛋白多态性-热力学参数
α-突触核蛋白等蛋白质的淀粉样原纤维是神经退行性疾病的标志,许多研究都集中在它们的动力学和形成机制上。关于这类结构的热力学稳定性的问题得到的关注要少得多,通过传统的技术方法也十分难以检测。
在传统研究蛋白聚集体热力学特征时,一般采用化学解聚的方法。化学解聚实验中使用的实验技术可分为两类:第一类通常使用的方法包括(超)离心或色谱(HPLC),但是十分耗时且样品消耗大。第二类技术包括圆二色性、光散射、自发荧光或硫黄素- t荧光。优点是分析混合物而不需要分离,从而实现快速和高通量的分析。然而,这些方法提供的是相对测量值,而不是所涉及物种的绝对浓度,这在分析过程中引入了一定程度的不确定性。此外,它们可能需要特定荧光团(固有荧光)的存在,并可能受到强散射(圆二色性)的限制。
但是在这个案例中,研究人员利用FIDA技术结合了以上两类技术的优点,研究了19种不同α-突触核蛋白原纤维多态性的热力学稳定性差异,并首次对这些差异进行了量化。并且证明了蛋白聚集体的形成可以在动力学或热力学控制下,通过溶液条件的变化可以稳定和破坏淀粉样原纤维。综上所述,我们的研究结果将热力学稳定性作为一个定义良好的关键参数,有助于更好地理解淀粉样纤维多态性的生理作用。
图6,不同结构α-syn蛋白热力学特征示意图
FIDA表征弥漫性和非弥漫性蛋白聚集体颗粒模型
脂质体、大纳米粒子、蛋白质聚集体)在实验过程中不能径向扩散(或在小于1分钟的实验时间尺度内边际扩散),并且在实验过程中保持在同一层流中,导致它们的信号在检测器处变形、不对称分布。在这里,我们利用这种现象来分离非弥散性聚集体和弥散性单体以确定淀粉样蛋白原纤维的稳定性。
首先用COMSOL 126软件(COMSOL Multiphysics®v. 6.1)对实验进行了数值模拟。
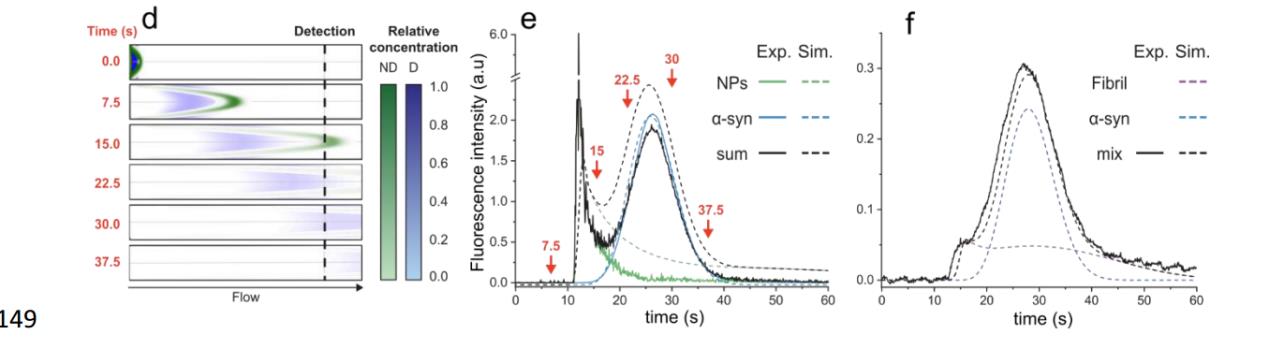
图7,为COMSOL 126软件模拟结果
为了验证我们的模拟,我们在FIDA仪器上使用αSyn 139单体(wt或F94W突变体)和荧光修饰的聚苯乙烯NPs (d = 200 nm;140 FluoSpheres™,Thermo Fisher)分别作为弥漫性和非弥漫性颗粒模型。两种物质的实验流动曲线几乎完全符合COMSOL模拟得到的。此外,非扩散粒子的FIDA数据与我们之前开发的Taylor色散诱导相分离(TDIPS )的解析十分近似。
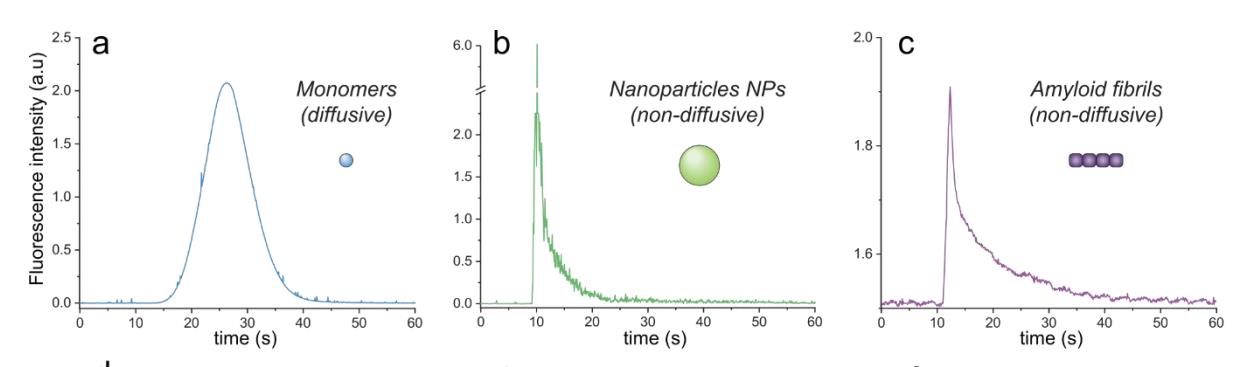 图8,为FIDA检测结果
图8,为FIDA检测结果
FIDA表征淀粉样蛋白原纤维的热力学稳定性与传统方法的对比
用不同的技术测量三种不同淀粉样蛋白原纤维的热力学稳定性。F94W αSyn突变原纤维的化学解聚用包括超速离心(UCF)、FIDA、nanoDSF、ThT和SLS在内的所有技术进行了研究。来自ThT荧光的数据略超出其他技术的误差范围。通过FIDA,ThT和SLS可以可靠地测量WT αSyn原纤维,但由于缺乏色氨酸(分子只含有4个酪氨酸),nanoDSF由于灵敏度的原因未能像预期的那样监测单体/原纤维的转化。PI3K-SH3淀粉样原纤维通过FIDA和nanoDSF测量可得到数值,但SLS强度数据不可靠,原因是在低变性剂浓度下发生沉淀。综述FIDA是唯一能够可靠测量所有3种模型的可靠技术。
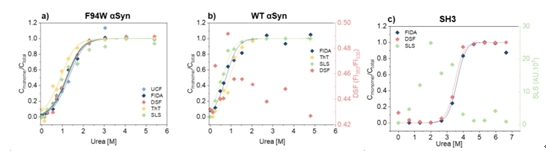 图9:用不同技术测定的三种不同淀粉样纤维的热力学稳定性。(a)采用超速离心法(UCF)、 FIDA、nanoDSF、ThT和SLS技术对F94W αSyn突变体纤维进行了化学解聚。ThT荧光测得的数据略超出其他技术的误差范围。(b) FIDA、ThT、SLS可以准确地检测WT αSyn纤维,然而,由于缺乏色氨酸,nanoDSF无法监测单体/纤维的变换。(c) FIDA和nanoDSF准确地测定了PI3K-SH3淀粉样纤维,但SLS强度数据不可靠,最可能的原因是在低变性剂浓度下发生沉淀
图9:用不同技术测定的三种不同淀粉样纤维的热力学稳定性。(a)采用超速离心法(UCF)、 FIDA、nanoDSF、ThT和SLS技术对F94W αSyn突变体纤维进行了化学解聚。ThT荧光测得的数据略超出其他技术的误差范围。(b) FIDA、ThT、SLS可以准确地检测WT αSyn纤维,然而,由于缺乏色氨酸,nanoDSF无法监测单体/纤维的变换。(c) FIDA和nanoDSF准确地测定了PI3K-SH3淀粉样纤维,但SLS强度数据不可靠,最可能的原因是在低变性剂浓度下发生沉淀
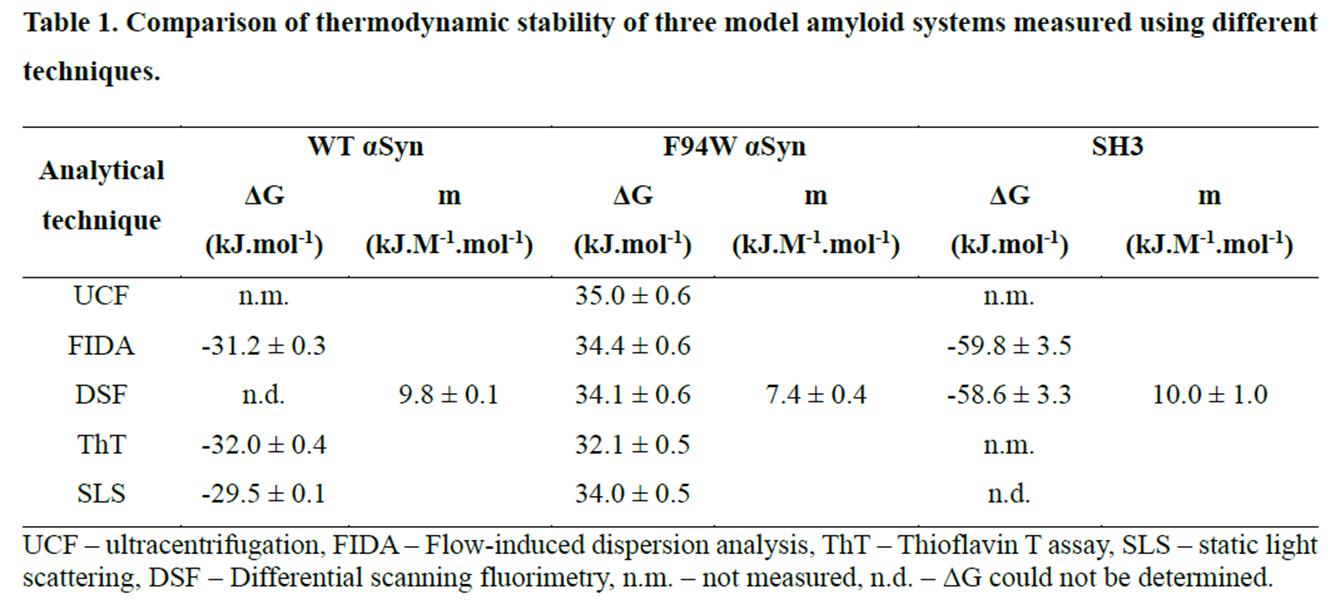
表1:比较各热稳定性测量方法:UCF -超速离心法,FIDA -流动诱导分散分析,ThT - Thioflavin T测定法,SLS -静态光散射法,DSF -差示扫描荧光法,n.m. -未测量,n.d. - ΔG不能确定
FIDA表征pH对纤维热稳定性的影响
在中性(pH 7.5)条件下和酸性(pH 5.5)条件下分别组装得到纤维Polymorph N和Polymorph H(图10. c)。Polymorph N在中性、Polymorhp H中性和酸性条件下,用不同浓度的尿素使其发生解聚(图10. b),Polymorph N和Polymohph H在各自对应的条件下,Polymorph H比 Polymorph N更稳定,但把Polymorph H放在中性条件下,它的稳定性大大下降,甚至比Polymorph N更不稳定(表2)。可以说,这种纤维稳定性的变化是由于αSyn在中性溶液中的溶解度高于酸性溶液,从而导致(部分)纤维在pH巨变时发生解聚。其他研究也观察到αSyn纤维在pH变化时发生类似的不稳定,这可能是体内发生的一个重要现象。
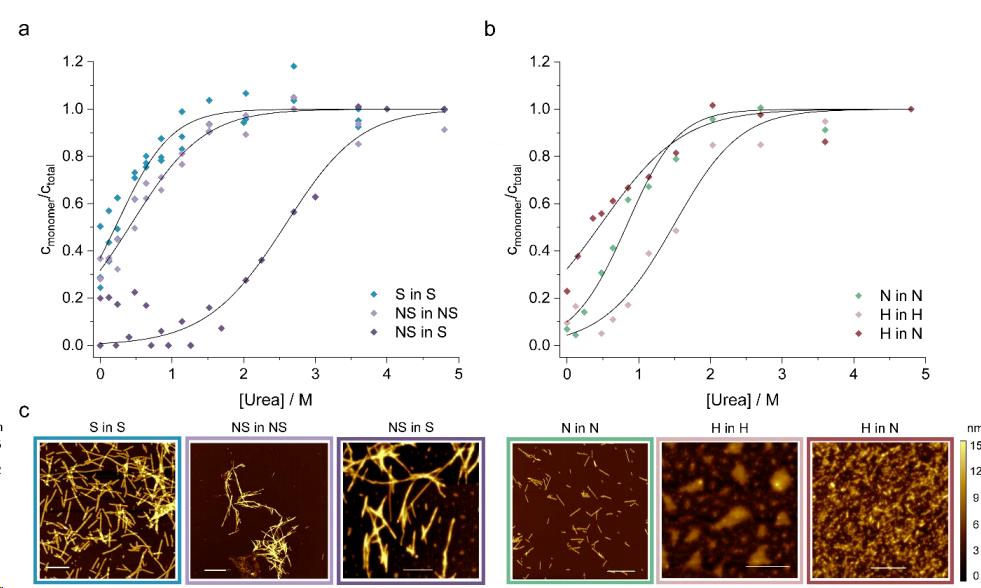
图10. 不同α - syn纤维在不同溶液条件下的解聚曲线。(a)在无盐(NS,紫色)或有盐(S,蓝色)的条件下制备和测量的纤维的稳定性。在盐(深紫色)的存在下进行解聚,以直接比较它们的稳定性。(b)在中性(pH 7.4, N,绿色)或酸性(pH 5, H,浅橙)条件下制备并测定纤维的稳定性。(c)AFM分析纤维形态
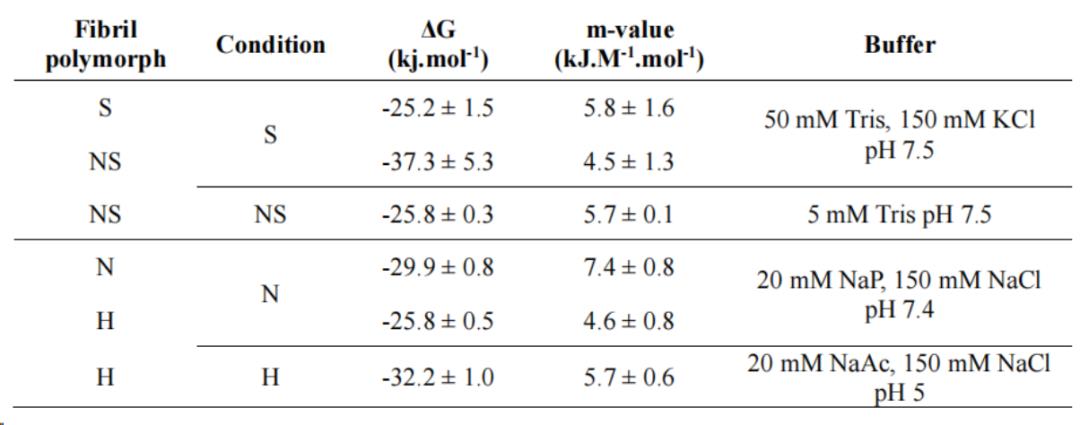
表2:在不同溶液条件下制备并用FIDA测定了不同WT αSyn纤维多态性的热力学稳定性。S-盐条件,NS-无盐条件,N-中性,H-酸性
来源:Farzadfard A, Kunka A, Mason TO, Larsen JA, Norrild RK, Torrescasana Dominguez E, et al. Thermodynamic characterization of amyloid polymorphism by Taylor dispersion analysis. ChemRxiv. 2023; doi:10.26434/chemrxiv-2023-41scv This content is a preprint and has not been peer-reviewed
利用FIDA确定抑制a-突触核蛋白寡聚物为靶点的小分子化合物作用机理
突触核蛋白(aSO)的小可溶性低聚物与神经元稳态的破坏有关,有助于帕金森病(PD)的发展。这使得aSO成为一个明显的药物靶点,但由于其丰度低、结构和形态复杂,开发有效的治疗方法受到挑战。
首先,利用数据库高通量筛选确定了四种神经元蛋白作为aSO相互作用的可能候选蛋白,即Cfl1、Uchl1、Sirt2和SerRS。
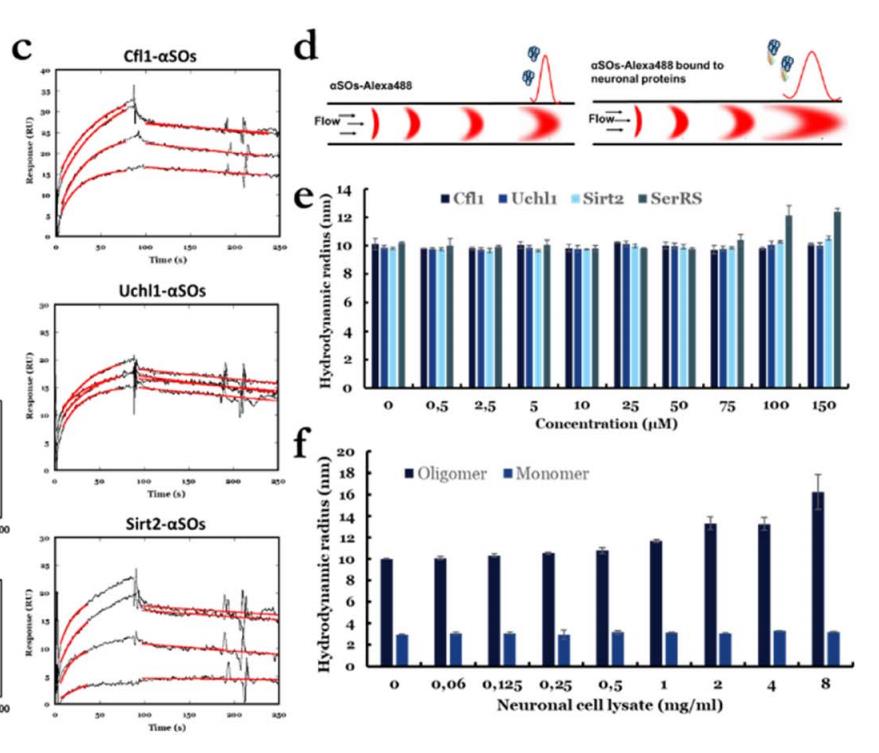
图11,(c) SPR分析测定的aSOs结合动力学。将1-15 mM的aSOs通过Cfl1、Uchl1和Sirt2固定在不同通道上的传感器表面。传感器图显示了SPR结合信号的时间曲线,其中使用1:1结合模型最适合结合区域的时间曲线以红色表示。(d)使用FIDA表征aSOs -配体相互作用的原理。(e) aSOs-Alexa488与蛋白配体结合后的Rh随配体浓度的变化。(f) aSOs-Alexa488和aSMs-Alexa488与不同浓度的神经元细胞裂解液结合后的Rh
将3种候选蛋白质分别固定在CM5芯片的通道上,测量不同aSO浓度下aSO的结合动力学(图c)。所有蛋白质都显示出结合信号的浓度依赖性增加(尽管Uch1的信号较弱),结果Cfl1、Uchl1和Sirt2的估计亲和力常数分别为0.29 μm、0.15 μm和0.6 μm。
但是蛋白质的在芯片的固定可能使结合数据的解释复杂化。因此,为了确定这些蛋白质在溶液中与aSOs的结合亲和力,研究者利用FIDA技术继续验证。
使用Alexa488标记的aSOs与未标记的配体组合作为指示剂。单独的aSO的Rh为10±0.2 nm,这与我们之前基于SEC-MALS和SAXS17报道的~11 nm的Rh相似,并验证了aSO的结构完整性。使用FIDAbio的PDB-Rh预测工具,我们预测了四个配体的Rh,它们分别为2.3nm(Cfl1)、2.4nm(Uchl1)、2.8nm(Sirt2)和3.6nm(SerRS)。在与配体结合后,aSO的Rh预计将增加到12–13 nm,并且这种增加完全在检测范围内(因为应该可以测量低至0.5 nm的增加)。然而,当用高达150mM C1、Uchl1和Sirt2滴定时,我们看到Rh几乎没有变化(图e)。只有SerRS的图的大小有一个小但显著的增加低聚物(从10到~12.5 nm),这仅在高浓度(100-150 mM)下发生。这一观察结果表明,配体与溶液中aSO的结合亲和力非常弱,远低于点印迹分析和SPR测量的亲和力。
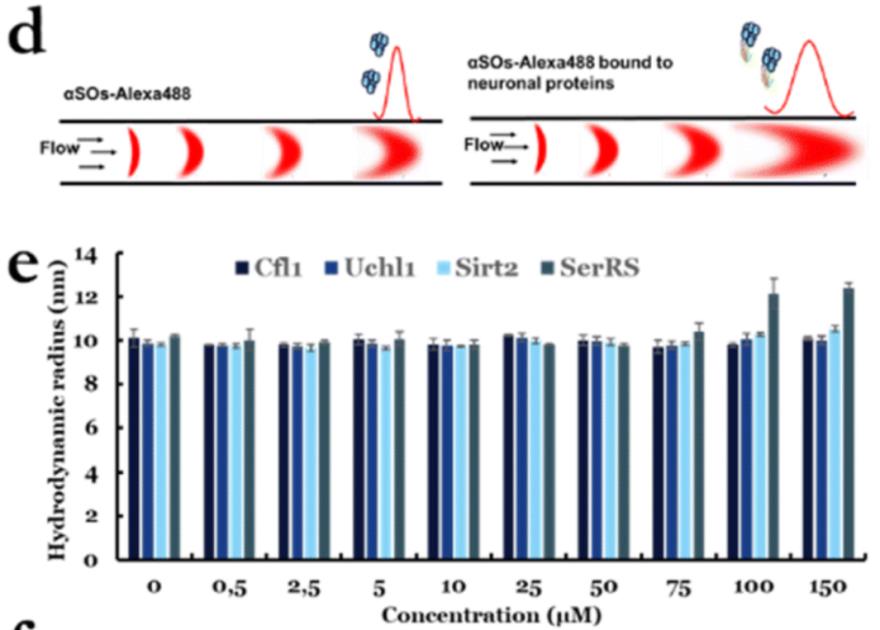
图12利用FIDA检测溶液中分子互作亲和力,配体与溶液中α-SOs的结合亲和力非常弱,远低于点印迹分析和SPR测量的亲和力
由于的候选蛋白与aSO的弱相互作用,研究者采用另一个思路使用aSO-膜系统来识别破坏aSO相互作用的小分子。
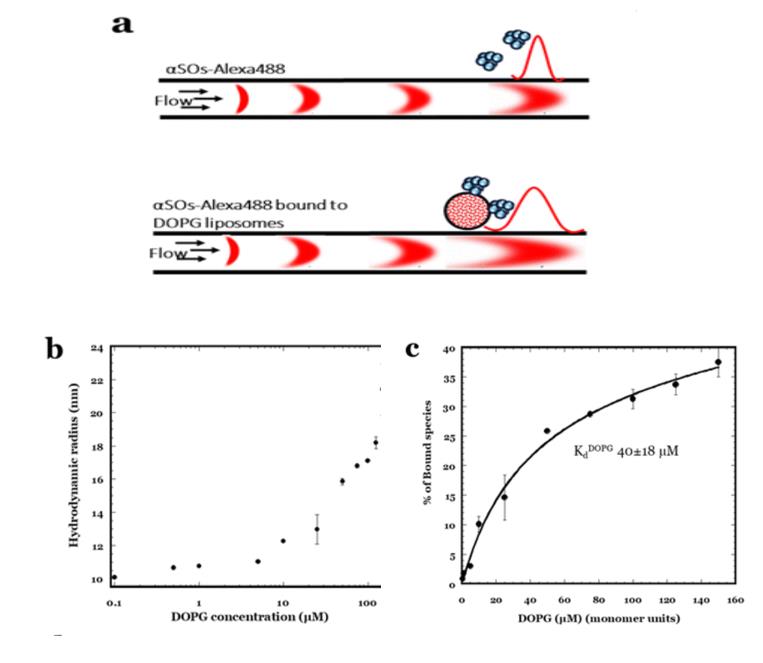
图13,aSOs-Alexa488与DOPG脂质体结合后的 Rh作为DOPG脂质浓度的函数。(c) 表观解离常数Kd(其中脂质浓度以单体单位表示)是使用来(b)的数据获得的
为了阐明与细胞膜结合的aSOs的细胞膜破坏性相互作用,制备了由DOPG脂质制成的模拟膜脂质体,并使用FIDA对其进行相互作用检测。在测量之前,将aSO-Alexa488和DOPG脂质体共孵育15分钟,随后在周围溶剂中用相应浓度的脂质体样品进行分析。
得到了aSOs-Alexa488与DOPG脂质体结合后的 Rh作为DOPG脂质浓度的函数。(c) 表观解离常数Kd(其中脂质浓度以单体单位表示)是使用来(b)的数据获得的
后面也用钙绿黄素的释放试验再次验证了,aSO蛋白对于膜的渗透性的增加,表面了蛋白与膜结合并破坏性造成膜通透性增加渗出钙离子的机制,验证了FIDA得出的结果。
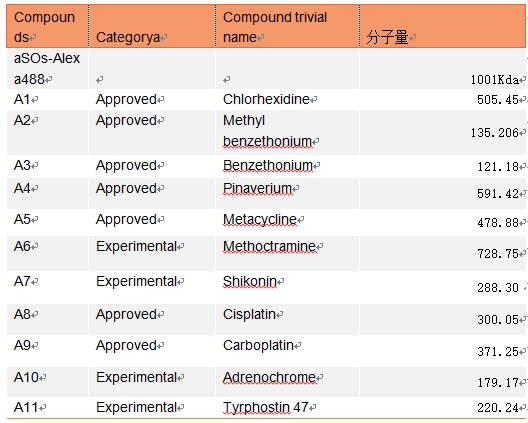
表3,筛选的11种化合物与aSOs-Alexa488分子量信息
为了寻找可破坏aSOs与膜结合的化合物,筛选了两个数据集的化合物,包括FDA批准的(Prestwick文库)和临床阶段的生物活性药物(Biomol文库)。我们使用EGCG和橄榄苦苷作为阳性对照和阴性对照,以钙绿黄素实验为指标筛选了11种最优化合物A1-A11。
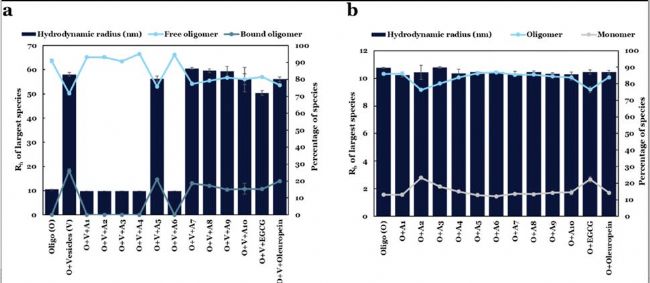
图14,在存在和不存在脂质体的情况下,通过FIDA分析αSO与化合物的相互作用。(a) 在DOPG脂质体和化合物存在下,最大种类的aSO-Alexa488的表观尺寸(Rh)。Rh的值和种类的百分比可以分别在左y轴和右y轴上读取。(b) 在化合物存在下的游离αSO-Alexa488(即不含脂质体)的Rh
综合a,b图与elisa结果,得出了小分子化合物结合抑制α核突触蛋白的三种作用机制:
1.A1-4,A6,完全抑制了膜复合体形成,但αSO大小不变
2.A5,7,8,9,10不完全抑制复合体形成
3.A2,3, EGCG增加了αSyn单体数量
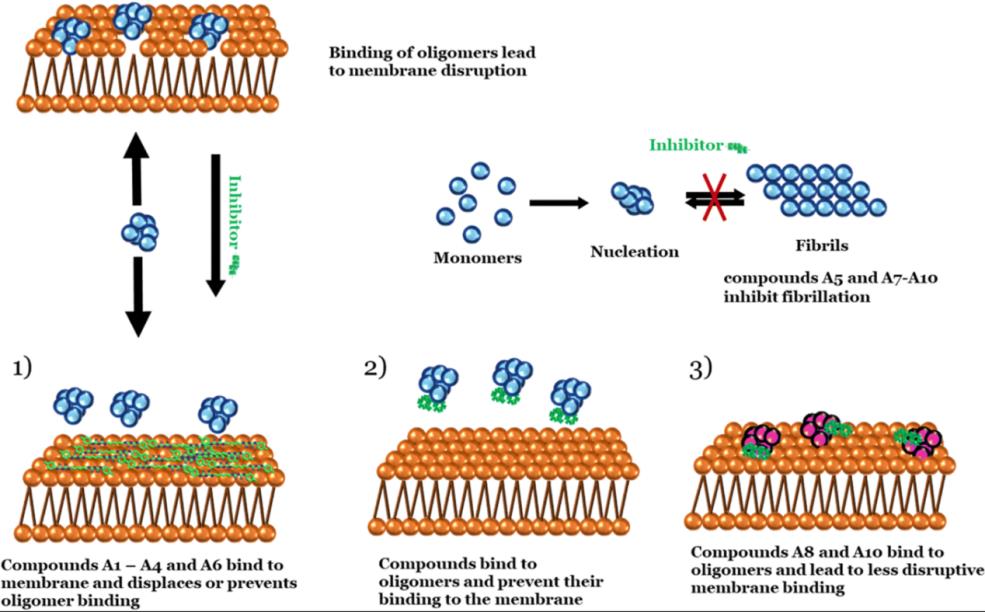
图15,化合物抑制α核突触蛋白的三种作用机制
来源:Drug repurposing screens identify compounds that inhibit α-synuclein oligomers' membrane disruption and block antibody interactions 2023
FIDA——Capflex方法表征α-突触核蛋白 (α-Syn) 液液相分离揭示淀粉样蛋白原纤维的形成机理
α-突触核蛋白 (α-Syn) 能够发生 LLPS ,随后可以由液相转变为固相,从而导致淀粉样蛋白原纤维的形成,这一过程可能是病理性的。
研究结果表明α-Syn液滴经历了液体到固体样的转变,这导致了含有纤维聚集体和低聚物的水凝胶的形成。此外,α-Syn甚至在细胞内形成液滴,随后转化为固体状聚集体,并且是由微管调节的。表明液液相分离是PD病理相关α-Syn聚集的第一步。
在100µM的α-Syn溶液中加入10nM的Alexa488标记的α-Syn与20% PEG6000,37℃孵育,每隔4小时(48小时)进行Capflex测量。在0h时,没有观察到峰值,这表明样品没有相分离;孵育4-8小时后,开始出现小而稀疏的峰,轻相浓度下降,这时相分离开始了;6h后,α-Syn相分离成液滴,稀相浓度下降到70µM,直到24h时几乎保持不变;当孵育时间达到30-48h时,即使在稀释样品后,信号峰值也没有消失(右图)。48h后,基线荧光达到2-5µM,表明α-Syn在指数相中发生了不可逆的液固转变和聚集。
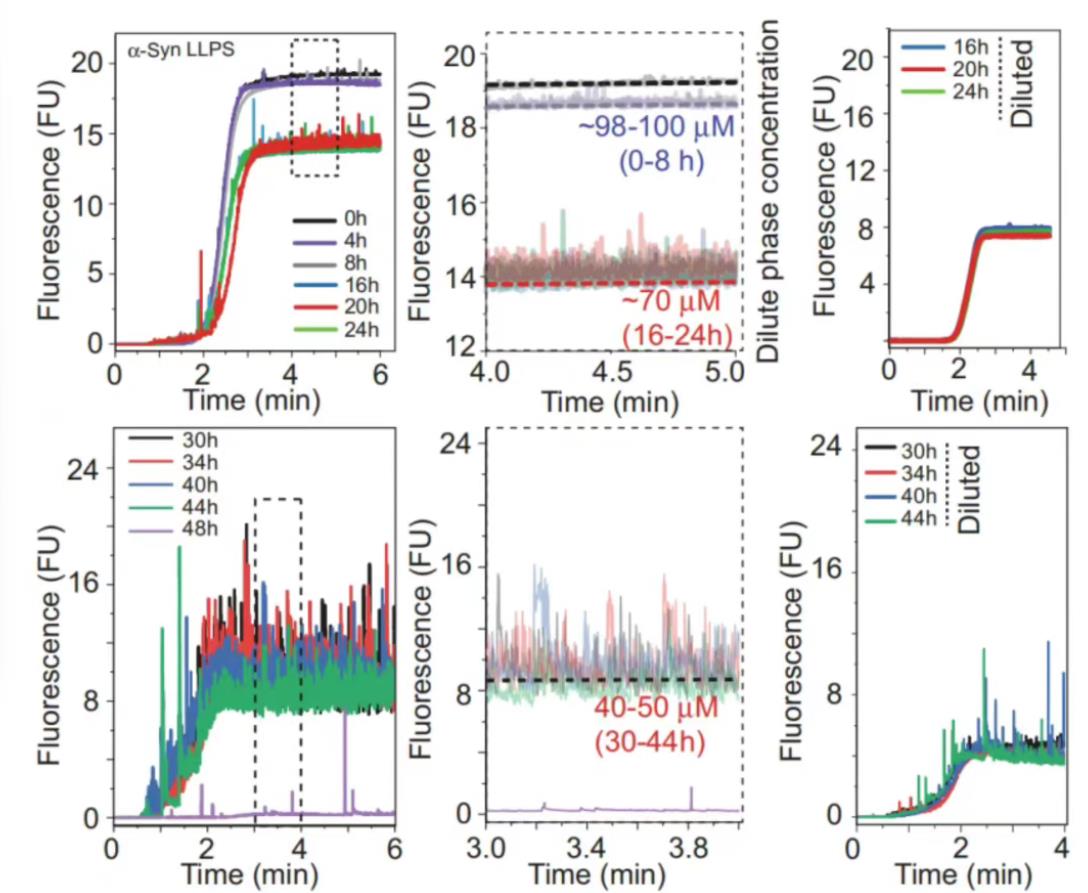
图16, 在100µM的α-Syn溶液中加入10nM的Alexa488标记的α-Syn与20%PEG6000,37℃孵育,每隔4小时(48小时)进行Capflex测量
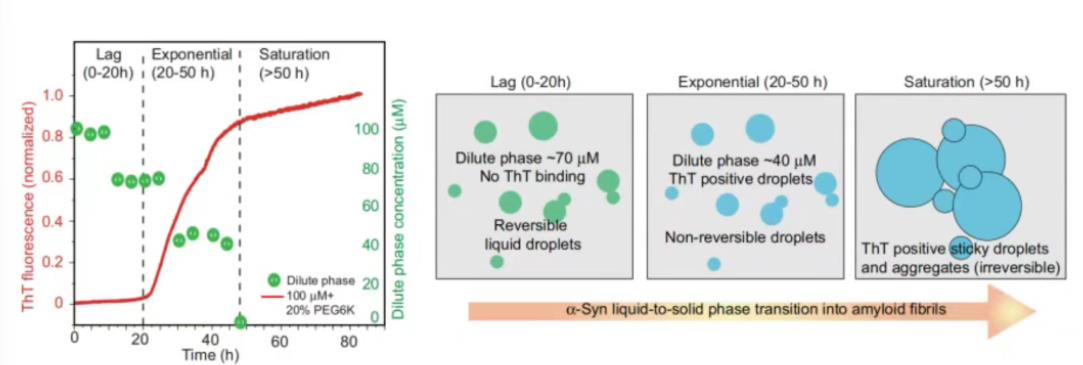
图17,ThT蛋白各阶段信号峰结果与α-Syn发生LLPS各阶段示意图
通过FIDA Aggregation分析,α-Syn LLPS溶液(100µM)发生LLPS的滞后时间为20-24小时;指数聚集阶段在孵育20-24小时后开始,并持续50小时到饱和(左图)。Capflex的数据清楚地表明,这种新的定量方法可检测到α-Syn发生LLPS的开始对应于蛋白聚集的滞后阶段。不可逆的液-固相变和淀粉样蛋白聚集主要发生在指数阶段(20-40h),这可以通过稀释相浓度的进一步降低和30h后的并且和ThT阳性信号峰结果一致得到证实。
FIDA定量表征LLPS的方法Capflex。与光漂白荧光恢复实验(FRAP)这类定性实验不同,其可以高通量、快速和准确地定量LLPS的多个关键参数:轻相浓度,相对液滴大小分布,以及液滴的形成,从而揭示淀粉样原纤维的动力学。
来源:Capillary flow experiments for thermodynamic and kinetic characterization of protein liquid-liquid phase separation,nature communications
FIDA检测FapA与FapB/FapC的相互作用机制
生物被膜(Biofilms)是指黏附于非生物或生物表面后,细菌通过分泌的胞外聚合物(extracellular polymeric substances, EPS)将自身包裹其中而形成的细菌聚集体膜状物。生物被膜对化学降解和抗生素具有高度的抗药性,使得这些感染频繁复发、治疗困难。生物膜具有高稳定性和抗性的原因之一是淀粉样原纤维,它能够稳固结构。靶向淀粉样蛋白形成、中断生物被膜形成是增加药物的抗菌性的有效策略。假单胞菌(Pseudomonas,Fap)的功能性淀粉样蛋白包括主要成分FapC、次要成分FapB(促进FapC成核)和小的周质蛋白FapA(条件原纤维的组成和形态),这三种成分的相互作用是Fap纤维发展的核心机制。为了更好的理解FapA影响原纤维的机制,作者利用FIDA研究了FapA与FapC/FapB 之间的相互作用。
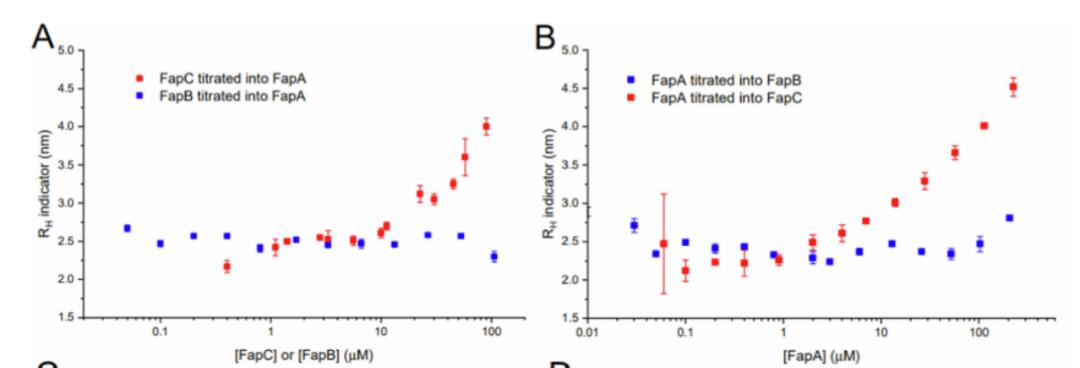 图18 .FIDA表征FapA与FapB/FapC相互作用。(A)1μM FapA-Alexa 488分别滴定FapC(红)、FapB(蓝);(B)0.25μM FapC-Alexa 488(红)、1.5μM-Alexa 488 FapB(蓝)分别滴定FapA
图18 .FIDA表征FapA与FapB/FapC相互作用。(A)1μM FapA-Alexa 488分别滴定FapC(红)、FapB(蓝);(B)0.25μM FapC-Alexa 488(红)、1.5μM-Alexa 488 FapB(蓝)分别滴定FapA
为了更好地了解FapA的潜在机制,使用FIDA研究了FapA与FapB或FapC之间的相互作用。用荧光探针Alexa Fluor 488标记FapA,并与逐渐增加的FapB或FapC预混液。FapA会在FapB或FapC之间形成络合物,当更多的FapB或FapC加入后,随着平衡向络合物形成时,分子水动力学半径大小应该增加。图(A)显示,加入FapC后,尺寸增加,而加入FapB不增加,说明FapA与FapC形成复合物,而不与FapB形成复合物。
来源:FapA is an Intrinsically Disorered Chaperone for Pseudomonas Functional Amyloid FapC 2023
关于FIDA

FIDA(Flow induced dispersion analysis,流动诱导分散分析)是最新推出的第一性原理的生物物理分析平台,利用泰勒分散的原理,基于粒子在层流状态下分散的特性,来测量粒子大小(流体力学半径Rh)。在层流状态下,流体在平行层中平滑地流动,不会发生混合。中心层流速要比贴近毛细管壁层的流速要快,从而产生了典型的抛物线流速度剖面。小的颗粒(如小分子\蛋白质)在层间扩散并以平均流速移动,使得它们在检测点的浓度呈高斯分布,它们的扩散系数(Dapp)可以通过拟合泰勒分散来获得,并根据Stokes-Einstein方程来确定它们的水动力半径(Rh)。这种技术符合第一原理定律,能够测量分子的真实大小Rh,具有非常重要的生物学意义,可以有效的监测蛋白早期的聚集(如Htt、Tau)、蛋白低聚物/纤维的相互作用。
对于较大的粒子(如脂质体、大纳米颗粒、蛋白聚集体等)在实验过程中不能呈径向扩散,且在实验过程中始终处于同一流层中,导致其在检测器上的信号分布不均匀。利用这种特性,可以从分散性的单体中区分出非分散性的聚合物(纤维),从而表征蛋白纤维的热稳定性 。不仅如此,FIDA是一项功能强大且全面的技术,从结构到结合再到热力学稳定性一应俱全。其测量的是分子的真实大小, FIDA的检测结果与现有方法具有一致性,同时实验成本极低,无需特殊的耗材,一次测量仅消耗40nL的样品;通过检测分子Rh的变化,对分子间的相互作用进行定性实验,操作简单,仅需几分钟就可以得到结果;通过滴定实验,以Rh的变化绘制结合曲线,可以得到分子互作的亲和力,从一个分子大小变化的角度去观察分子间的相互作用,同时还可以通过构象导致的分子荧光变化来获得亲和力,对于亲和力的获取可以同时用正交的方法验证。此外,FIDA可以测定Kon/Koff,对于分子与聚集体的结合动力学可以有效表征。
FIDA技术检测速度快,无需担心样品在检测期间发生变化从而影响结果。检测灵敏度高,不仅仅依赖于单一指标,可多维表征蛋白聚集体粒径,结构,亲和力,动力学与热力学,并且还能反馈样品聚集信息。不同类型的蛋白聚集体样本都可通过FIDA进行可靠检测,这个功能是其他技术无法实现的。
FIDA在神经退行性疾病研究的相关文献:
1.Thermodynamic characterization of amyloid polymorphism by microfluidic transient incomplete separation 2023
2.Thermodynamic characterization of amyloid polymorphism by Taylor dispersion analysis 2023
3.Specific inhibition of α-synuclein oligomer generation and toxicity by the chaperone domain Bri2 BRICHOS 2023
4.Fibril Paint to detect amyloids and determine fibril length 2023
5.Drug repurposing, inhibit aSN oligomers' membrane disruption 2023
6.FapA is an Intrinsically Disorered Chaperone for Pseudomonas Functional Amyloid FapC 2023
7.Capillary flow experiments for thermodynamic and kinetic characterization of protein liquid-liquid phase separation 2021
8.Drug repurposing screens identify compounds that inhibit α-synuclein oligomers' membrane disruption and block antibody interactions 2023
